咸海龙属
-
所属目录:
自然科学
-
更新时间:
2023-09-07 16:59
简介
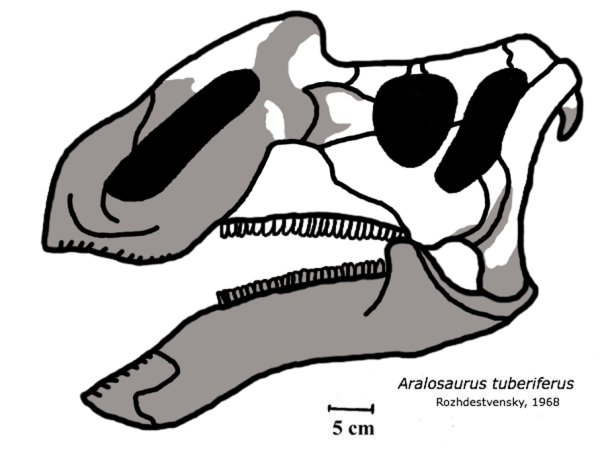
咸海龙属化石时期:白垩纪后期, 83.6 Ma PreЄЄOSDCPTJKPgN↓咸海龙的旧版头骨重建图,白色区块为已发现部位;在这个版本中咸海龙被错误地诠释为类似格里芬龙的鸭嘴龙亚科。科学分类 界:动物界 Animalia门:脊索动物门 Chordata纲:蜥形纲 Sauropsida总目:恐龙总目 Dinosauria目:†鸟臀目 Ornithischia亚目:†鸟脚亚目 Ornithopoda科:†鸭嘴龙科 Hadrosauridae亚科:†赖氏龙亚科 Lambeosaurinae族:†咸海龙族 Aralosaurini属:†咸海龙属 AralosaurusRozhdestvensky, 1968模式种突吻咸海龙Aralosaurus tuberiferusRozhdestvensky, 1968咸海龙(属名:Aralosaurus)是鸭嘴龙科恐龙的一个属,生活于白垩纪晚期、相当于现在的哈萨克。它的化石只有头骨后半部(缺少下颌)以及一些颅后骨骼,发现于哈萨克中部的波斯妥贝组(英语:Bostobe Formation),年代可追溯至上桑托阶-下坎潘阶交界,约8360万年前。在附近也发现了其他近亲,例如牙克煞龙。模式种也是唯一已知的物种突吻咸海龙(Aralosaurus tuberiferus)于1968年由阿纳托利·康斯坦丁诺维奇·罗特杰斯特文斯基所叙述、命名。属名意为“来自咸海的蜥蜴”,因化石发现于咸海东北部地区;种名tuberiferus意指“长着有如块茎般隆起”,因在化石鼻骨后部有明显的衍生突起,到眼眶前大幅升高。咸海龙起初被重建为具有类似北美小贵族龙(根据与现在被归于格里芬龙标本进行比较)的鼻弓,于是长期以来都被分类于鸭嘴龙亚科底下,这个分类直到2004年被推翻,因为在对头骨重新检验后,足以鉴定出许多赖氏龙亚科的典型特征。值得一提的是,这项研究中指出,咸海龙眼眶前面远处有个中空骨质结构,与呼吸管相连。该结构从底部碎裂了,很不幸地其形状与尺寸不得而知。在更近期的研究,咸海龙被鉴定为最基础的赖氏龙亚科,并与近缘的似鸭龙(法国晚马斯特里赫特阶)一同分类于新建立的演化支咸海龙族。发现及研究历史
正模标本(PIN 2229,目前存放于俄罗斯科学研究院的古生物部门)于1957年由苏联考察队在哈萨克(当时属于苏联一部分)Shakh-Shakh地点附近所发现。所知并不多,主要有一个不完整的部分头骨,缺乏大部分吻部与整个下颌。除此之外还有发现单独的牙齿及一些破碎的颅后骨骼,包含尺骨、桡骨、股骨、胫骨、腓骨、距骨、跖骨,其中唯一完整的是一个肱骨和一个跖骨。1968年,苏联古生物学家阿纳托利·康斯坦丁诺维奇·罗特杰斯特文斯基((Анатолий Константинович Рождественский))叙述、命名了模式种突吻咸海龙(Aralosaurus tuberiferus)。他估计头骨全长65公分、身体全长约6.5米。根据颅部缝线尚未愈合,推测属于一只未成熟个体。在当时罗特杰斯特文斯基认为咸海龙的头骨眼眶前具有鼻弓,而与小贵族龙最为相似;但当时作比较的标本现在已重新归在格里芬龙,而非小贵族龙,虽然它们彼此近缘。于是咸海龙被分类在不具有头冠的鸭嘴龙亚科。
2004年,帕斯卡·古德弗里特(Pascal Godefroit)和同事重新检验了咸海龙头骨,根据他们所述,原先由罗特杰斯特文斯基鉴定为鼻弓中部和前上颌骨后部的碎片似乎遗失了。这些碎片最初是单独找到的,而头骨其余部分关节相连(包含鼻骨后部碎片)。于是现在不再能够确认存在着鼻弓。另一方面,此研究表示咸海龙并非是种类似格里芬龙的鸭嘴龙科,因为所谓的鼻冠实际上是中空结构的一部分,且他们并未在咸海龙身上发现任何鸭嘴龙亚科的鉴定特征。相反地,咸海龙头骨展现了几个特征(上颌骨、顶骨、鳞状骨、特别是鼻骨)得出它应该属于赖氏龙亚科的结论。值得注意的是,明显与其余头骨关节相连的鼻骨后部碎片在眼眶正前方隆起,并参与构成中空头冠结构。鉴于此结构已从底部碎裂,无法得知其尺寸及形状。但从吻部视角来看,鼻骨很明显是中空地,并由一道直接与鼻骨后腹侧相通的宽凹槽深凿而出。很明显的咸海龙的头冠环绕着呼吸系统的一部分。中空头冠内部的鼻腔延伸是赖氏龙亚科的典型特征,于是确定咸海龙属于赖氏龙亚科。咸海龙亦展现数项头骨特征代表它是赖氏龙亚科的基础物种:虽然具有中空鼻腔结构,位置却在眼眶之前,属于赖氏龙亚科的原始情况;在最进阶的物种中,因为前上颌骨的重大发育,使鼻骨后移到头部尾背侧;此外,在较衍化的赖氏龙亚科中,头盖骨产生变异来构成中空头冠的固定位置,但在咸海龙中,头盖骨没有发生任何变异。不幸的是,原本被鉴定为鼻弓部位碎片已经遗失,本来可以依照这些碎片尝试重现咸海龙的头部轮廓。因此还需要发现更完整的化石才能了解关于中空头冠的详细资讯。
描述
咸海龙的体型就像一头象,全长约 6.5 至 9 米,体重可能相当于一头犀。前面喙嘴无齿,用来切下植物,并在嘴巴后部有着 30 列牙齿构成的齿系,总计约 1000 颗,用来咀嚼,在草食性鸟臀目非常普遍,但却在爬行动物相当少见。头骨背侧很宽,显示有大型颌部肌肉可供咀嚼。眼睛前有着低矮的隆起物,亦常见于大部分鸭嘴龙科。它们可能会吹气使其如气球般膨胀,以充当声音放大器,可能可以用来作为视觉展示、吓阻掠食者、召集族群或吸引异性。虽然还没有确定的身体骨骼证据,推测它也有着鸭嘴龙科的标准共有特征:粗大尾巴、巨大身躯及有力后脚。
分类
早期的研究将咸海龙归类于鸭嘴龙科下的鸭嘴龙亚科,这是一群没有头冠的演化支;因为当时认为巴克龙、冠长鼻龙、小贵族龙的头骨结构与咸海龙相似。
直到2013年鸭嘴龙类专家亚伯特·皮耶妥-马奎兹(Albert Prieto-Marquez)等人发表似鸭龙属的论文,对整个赖氏龙亚科恐龙的多样性、生物地理及其分类系统做了全面性分析。他们将咸海龙重新分类到赖氏龙亚科底下,并是整个家族中最原始的物种;同时也认为咸海龙与似鸭龙互成姊妹群,于是在赖氏龙亚科基部新建立了一个演化支咸海龙族(Aralosaurini)来包含这两属。以下演化树取自其研究:
赖氏龙亚科 Lambeosaurinae
咸海龙族 Aralosaurini
咸海龙属 Aralosaurus
似鸭龙属 Canardia
牙克煞龙属 Jaxartosaurus
青岛龙族 Tsintaosaurini
青岛龙属 Tsintaosaurus
似凹齿龙属 Pararhabdodon
副栉龙族 Parasaurolophini
卡戎龙属 Charonosaurus
副栉龙属 Parasaurolophus
短冠副栉龙 Parasaurolophus cyrtocristatus
小号手副栉龙 Parasaurolophus tubicen
沃克氏副栉龙 Parasaurolophus walkeri
赖氏龙族 Lambeosaurini赖氏龙属 Lambeosaurus
赖氏赖氏龙 Lambeosaurus lambei
大冠赖氏龙 Lambeosaurus *gnicristatus
冠龙属 Corythosaurus
卡萨鲁斯冠龙 Corythosaurus casuarius
间型冠龙 Corythosaurus intermedius
斯氏“亚冠龙” *Hypacrosaurus* stebingeri
亚冠龙属 Hypacrosaurus
大天鹅龙属 Olorotitan
艾瑞龙属 Arenysaurus
布拉西龙属 Blasisaurus
巨保罗龙属 Magnapaulia
威拉弗龙属 Velafrons
阿穆尔龙属 Amurosaurus
黑龙江龙属 Sahaliyania
但尼克·朗里奇(Nicholas Longrich)2020年发表异客龙的研究中,对赖氏龙亚科的分类提出了不同看法。咸海龙仍位于最基础位置,并与其他亚洲物种(牙克煞龙、日本龙、青岛龙)、另一个包含欧洲及非洲物种的演化支艾瑞龙族(Arenysaurini)有关联:
鸭嘴龙科 Hadrosauridae
栉龙亚科 Saurolophinae
赖氏龙亚科 Lambeosaurinae
咸海龙 Aralosaurus
牙克煞龙 Jaxartosaurus
日本龙 Nipponosaurus
青岛龙 Tsintaosaurus
艾瑞龙族 Arenysaurini
艾瑞龙 Arenysaurus
似凹齿龙 Pararhabdodon
匙龙 Koutalisaurus
巴斯妥斯的赖氏龙亚科(英语:Basturs Poble bonebed) Basturs lambeosaurine
似鸭龙属 Canardia
弱肩龙 Adynomosaurus
瑟拉德罗斯提尔的赖氏龙亚科 Serrat del Rostiar lambeosaurine
布拉西龙 Blasisaurus
异客龙 Ajnabia
副栉龙族 Parasaurolophini
赖氏龙族 Lambeosaurini
古生物学
古生物地理学
咸海龙生存于近缘物种似鸭龙之前2000万年。后者生活在伊比利-阿莫里凯(英语:Armorica)群岛,是白垩纪晚期欧洲群岛最西侧的岛屿。因为在伊比利-阿莫里凯群岛晚坎潘阶-早马斯特里赫特阶盛产化石的地方都还没发现过赖氏龙亚科,推测咸海龙的后裔应该在更晚的时间才抵达这里,估计是早马斯特里赫特阶之末或晚马斯特里赫特阶。然而,后来西班牙发现数种赖氏龙亚科遗骸,显示它们在早马斯特里赫特阶已达到最高多样性,从而反驳了前面的理论。这段物种迁徙可能经历了几个过程,首先为西亚大陆和东欧群岛间暂时性的陆地连接,再来是不同欧洲诸岛到伊比利-阿莫里凯群岛之间移动。似鸭龙在当时当地是相当原始的物种,暗示了伊比利-阿莫里凯群岛可能是咸海龙族的最后幸存地。
古生态学
波斯妥贝组(英语:Bostobe For*tion)沉积层主要由黏土和砂岩构成,并呈现当时为泛滥平原至河口湾的环境,属于热带或副热带气候。这些生物群落位于当时的亚洲大陆西海岸、古土尔介海(英语:Turgai Sea)边缘,后者与特提斯洋及北冰洋接壤。由于来自大陆上升气流引起的强风条件,在这片沿岸冲积平原以西的浅海是个生物丰沛的产地。强风也是造成桑托阶至早坎潘阶当地气候明显干旱的原因,使被子植物群落产生变化,大型阔叶型植物显得稀少罕见,而小型窄叶型的榆科占据主导地位。被子植物占整个植物群落的75%,其余则是松柏门、罕见的银杏及苏铁。
发现咸海龙的Shakh Shakh地区盛产高多样性的动物群,包含:许多鱼类(软骨鱼纲、软骨硬鳞鱼、全骨下纲)、青蛙、至少六种乌龟(有些壳长达75公分)、石龙子、鳄鱼、翼龙类(咸海神翼龙)、鸟类、哺乳类、以及数种未定的恐龙(甲龙科、蜥脚类、暴龙超科、似鸟龙类、镰刀龙类、伤齿龙科、驰龙科)。邻近的Akkurgan地区鉴别出至少两种鸭嘴龙科(阿斯坦龙、勇士龙);翼龙类的魔鸟翼龙(英语:Samrukia)(下颌一度被认为属于偷蛋龙类的近颌龙科、再来是种大鸟),可能是咸海神翼龙的次异名。
延伸阅读
鸭嘴龙类研究历史(英语:Timeline of hadrosaur research)
参考来源
^ 1.0 1.1 1.2 1.3 Godefroit, P.; Alifanov, V.; Boltsky, Y. A re-appraisal of *Aralosaurus tuberiferus* (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre. 2004, (74): 139–154 . (原始内容存档于2021-05-23).
^ 2.0 2.1 2.2 2.3 2.4 2.5 2.6 Rozhdestvensky, A.K. Gadrozavry Kazakhstana . (PDF). Akademia Nauk SSSR, Moscow. 1968: 97–141 . (原始内容存档 (PDF)于2014-08-19).
^ 3.0 3.1 3.2 Holtz, Tho*s R. Jr. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. 2012. p.46. Winter 2011 Appendix (页面存档备份,存于互联网档案馆).
^ 4.0 4.1 4.2 4.3 4.4 Prieto-Márquez, A.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À. Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis. PLOS ONE. 2013, 8 (7): e69835. Bibcode:201*LoSO...869835P. PMC 3724916 . PMID 23922815. doi:10.1371/journal.pone.0069835.
^ 5.0 5.1 Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A.; Gillette, David D.; Norell, Mark A.; Olshevsky, George; Parrish, J. Michael & Weishampel, David B. *Aralosaurus*. The Age of Dinosaurs. Publications International, LTD. 1994: p.126. ISBN 0-7853-0443-6. 引文格式1维护:冗余文本 (link)
^ Longrich, Nicholas R.; Suberbiola, Xabier Pereda; Pyron, R. Alexander; Jalil, Nour-Eddine. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretaceous Research. 2020, 120: 104678 . doi:10.1016/j.cretres.2020.104678. (原始内容存档于2021-04-04).
^ 7.0 7.1 Csiki-Sava, Z.; Buffetaut, E.; Ősi, A.; Pereda-Suberbiola, X.; Brusatte, S.L. Island life in the Cretaceous-faunal composition, biostratigraphy, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. ZooKeys. 2015, (469): 1–161. PMC 4296572 . PMID 25610343. doi:10.3897/zookeys.469.8439.
^ Fondevilla, V.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À.; Moncunill-Solé, B.; Köhler, M. Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) re*ins from Basturs Poble bonebed (late early Maastrichtian, tremp Syncline, Spain). PLOS ONE. 2018, 13 (10): e0206287. PMC 6209292 . PMID 30379888. doi:10.1371/journal.pone.0206287.
^ Prieto-Márquez, A.; Fondevilla, V.; Sellés, A.G.; Wagner, J.R.; Galobart, À. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago. Cretaceous Research. 2019, 96: 19–37. doi:10.1016/j.cretres.2018.12.002.
^ 10.00 10.01 10.02 10.03 10.04 10.05 10.06 10.07 10.08 10.09 10.10 10.11 10.12 Averianov, A.; Dyke, G.; Danilov, I.; Skutschas, P. The paleoenvironments of azhdarchid pterosaurs localities in the Late Cretaceous of Kazakhstan. ZooKeys. 2015, (483): 59–80. PMC 4351447 . PMID 25755624. doi:10.3897/zookeys.483.9058.
^ Danilov, I.G.; Vitek, N.S.; Averianov, A.O.; Glinskiy, V.N. A new soft-shelled trionychid turtle of the genus Khunnuchelys from the Upper Cretaceous Bostobe For*tion of Kazakhstan. Acta Palaeontologica Polonica. 2015, 60 (1): 155–161. doi:10.4202/app.2013.0045 .
^ Averianov, A.O. New records of azhdarchids (Pterosauria, Azhdarchidae) from the Late Cretaceous of Russia, Kazakhstan, and Central Asia. Paleontological Journal. 2007, 41 (2): *–197. doi:10.1134/S0031030107020098.
^ Dyke, G.J.; Malakhov, D.V. Abundance and taphonomy of dinosaur teeth and other vertebrate re*ins from the Bostobynskaya For*tion, north-east Aral Sea region, Republic of Kazakhstan. Cretaceous Research. 2004, 25 (5): 669–674. doi:10.1016/j.cretres.2004.06.004.
^ Averianov, A.O. Theropod dinosaurs from Late Cretaceous depositsin the northeastern Aral Sea region, Kazakhstan. Cretaceous Research. 2007, 28 (3): 532–544. doi:10.1016/j.cretres.2006.08.008.
^ Averianov, A. Frontals bones of non-avian theropod dinosaurs from the Upper Cretaceous (Santonian-?Campanian) Bostobe For*tion of the northeastern Aral Sea region, Kazakhstan. Canadian Journal of Earth Sciences. 2015, 53 (2): 168–175. doi:10.1139/cjes-2015-0099.
^ Naish, D.; Dyke, G.; Cau, A.; Escuillié, F.; Godefroit, P. A gigantic bird from the Upper Cretaceous of Central Asia. Biology Letters. 2012, 8 (1): 97–100. PMC 3259976 . PMID 21835881. doi:10.1098/r*l.2011.0683.
^ Buffetaut, E. Samrukia nessovi, from the Late Cretaceous of Kazakhstan: A large pterosaur, not a giant bird. Annales de Paléontologie. 2011, 97 (3–4): 133–138. doi:10.1016/j.annpal.2011.10.001.
查
论
编
鸟脚亚目界: 动物界
门: 脊索动物门
演化支:蜥形纲
总目: 恐龙总目
目: 鸟臀目
参见 真爬行动物
参见 主龙形下纲
参见 鸟臀目
见下文↓
脚
亚
目
布氏龙属
长眠龙属
群聚龙属
吉迪恩曼特尔龙属(英语:Gideon*ntellia)?
棱齿龙属?
韦瓦拉龙属
阿纳拜斯龙属
阿特拉斯科普柯龙属
洪奔龙属
加利恩龙属
加斯帕里尼龙属
伊萨西奔龙属
巨谜龙属
岭奔龙属
莫罗龙属?
南方棱齿龙属?
快达龙属
岛屿龙属?
小头龙属
特立尼龙属
龙
类
腱龙属
近龙属?
福斯特龙属?
木他龙属
马氏齿龙属(英语:Matheronodon)
栅齿龙属
入侵者龙属
凹齿龙属
特兰西瓦尼亚龙属
查摩西斯龙属
树
龙
形
类(英语:Dryomorpha)
卡洛夫龙属
橡树龙属
难捕龙属
艾尔雷兹龙属
东橡树龙属
稚龙属?
康纳龙属
旷野龙属
拇
指
龙
类
弯龙属
龙爪龙属
欧文齿龙属
里氏鸭龙属
棘
龙
类
重骨龙属
巴彦淖尔龙属
雪松山龙属
库姆纳龙属
达科塔齿龙属
福井龙属
马龙属
高刺龙属
巨禽龙属
兰州龙属
沉龙属
巨手龙属(英语:Magna*nus)
那派龙属
峡谷龙属(英语:O*akasaurus)
扁臀龙属
船首龙属
众神花园龙属
犹他齿龙属
呵叻龙属(英语:Ratchasi*saurus)?
薄氏龙属
布赖斯通龙属
锦州龙属
曼特尔龙属
莫雷利亚齿龙属
豪勇龙属
禽龙属
嘴
龙
超
科
高吻龙属
勇士龙属
乔伊尔齿龙属
大同龙属
马鬃龙属
公婆泉龙属
高志龙属
鸭颌龙属
原巴克龙属
暹罗齿龙属
诗琳通龙属
叙五龙属
左云龙属
波尔特利龙属
嘴
龙
形
态
类
巴克龙属
破碎龙属
原赖氏龙属
戍卫龙属
计氏龙属
戈壁鸭龙属
古鸭龙属
研颌龙属
金塔龙属
列弗尼斯氏龙属
冠长鼻龙属
南宁龙属
南阳龙属
近鸭嘴龙属
始鸭嘴龙属
双庙龙属
谭氏龙属
沼泽龙属
特提斯鸭嘴龙属
云冈龙属
张衡龙属
嘴
龙
科
鹰鼻龙属
始糙齿龙属
鸭嘴龙属
拉潘帕龙属
大和龙属
女巫龙属
鸭
嘴
龙
类
氏
龙
亚
科
阿穆尔龙属
咸海龙属
牙克煞龙属
哈萨克赖氏龙属
日本龙属
弱肩龙属
异客龙属
艾瑞龙属
布拉西龙属
似鸭龙属
似凹齿龙属?
冠龙属
亚冠龙属
赖氏龙属
巨保罗龙属
扇冠大天鹅龙属
黑龙江龙属
帆额龙属
疑冠龙属
弯嚼龙属?
卡戎龙属
副栉龙属
符栉龙属
似凹齿龙属?
青岛龙属
龙
亚
科
巴思钵氏龙属
始无冠龙属
短冠龙属
慈母龙属
饰面龙属
原短冠龙属
乌拉嘎龙属
埃德蒙顿龙属
神威龙属
冥犬龙属
昆杜尔龙属
莱阳龙属
山东龙属
古植食龙属?
阿纳萨齐龙属
波拿巴龙属?
钩鼻龙属
鹜龙属
赤原龙属
分离龙属
宽鼻龙属
纳秀毕吐龙属
鼻王龙属
独孤龙属
南似鸭龙属?
奥氏栉龙属
波拿巴龙属?
原栉龙属
栉龙属
钝指龙足迹(英语:Amblydactylus)
异样龙足迹(英语:Anomoepus)
弯龙足迹
卡利尔足迹(英语:Caririchnium)
达科塔龙(英语:Dakotasaurus)
纳瓦霍足迹(英语:Dinehichnus)
鸭嘴龙足迹(英语:Hadrosauropodus)
禽龙足迹
嘉荫龙足迹(俄语:Jiayinosauropus)
Orcauichnites(英语:Orcauichnites)
鸟脚龙足迹(英语:Ornithopodichnites)
Shiraminesauropus(英语:Shiraminesauropus)
Sousaichnium(英语:Sousaichnium)
Staurichnium(英语:Staurichnium)
四足龙足迹(英语:Tetrapodosaurus)
温顿足迹(英语:Wintonopus)
Guegoolithus(英语:Guegoolithus)
阿斯坦龙属
柱齿龙
碎嘴龙属?
双芽龙属
优尾龙属?
泥隐龙属
酋长龙属
满洲龙属
小鸭嘴龙属
正骨龙属
帆骨盆龙属
呵叻龙属(英语:Ratchasi*saurus)
里亚宾宁鸭嘴龙属(英语:Riabininohadros)
史蒂芬龙属(英语:Stephanosaurus)
强龙属
墙骨龙属
糙齿龙属
亚琛龙属
阿尔哈拉路龙属
鸭嘴龙类的食性与进食方式
棱齿龙科
埃德蒙顿龙木乃伊 AMNH 5060
埃德蒙顿龙木乃伊 S.M. R4036
南似鸭龙属
数据: Q132769
EoL: 10571841
Fossilworks: 38769
GBIF: 4823273
IRMNG: 1278683
生物学主题
恐龙主题
取自“https://zh.*.org/w/index.php?title=鹹海龙属&oldid=70930096”